
Nature子刊:许琛琦/施小山团队阐述免疫受体的近膜静电调控理论
来源:生物世界 2024-11-24 16:47
该文章明确定义了BRS信号基序,总结了其近膜信号转导机制,阐述了免疫受体BRS突变与人类疾病的相关性,并探讨了BRS在创新免疫疗法中的应用潜力。
中国科学院分子细胞科学卓越创新中心许琛琦、中国科学院深圳先进技术研究院合成生物学研究所施小山等在 Nature 子刊 Nature Reviews Immunology 上发表了题为Charge-based immunoreceptor signalling in health and disease 的展望文章,深入探讨了一类普遍存在的信号基序-碱性残基富集序列(basic-residue-rich sequence,BRS)。跨膜蛋白质中存在近膜正电残基,是经典生化教科书中的已知结论。但是这些序列的信号机制,生理病理功能和应用前景却没有系统性总结。该文章明确定义了BRS信号基序,总结了其近膜信号转导机制,阐述了免疫受体BRS突变与人类疾病的相关性,并探讨了BRS在创新免疫疗法中的应用潜力。

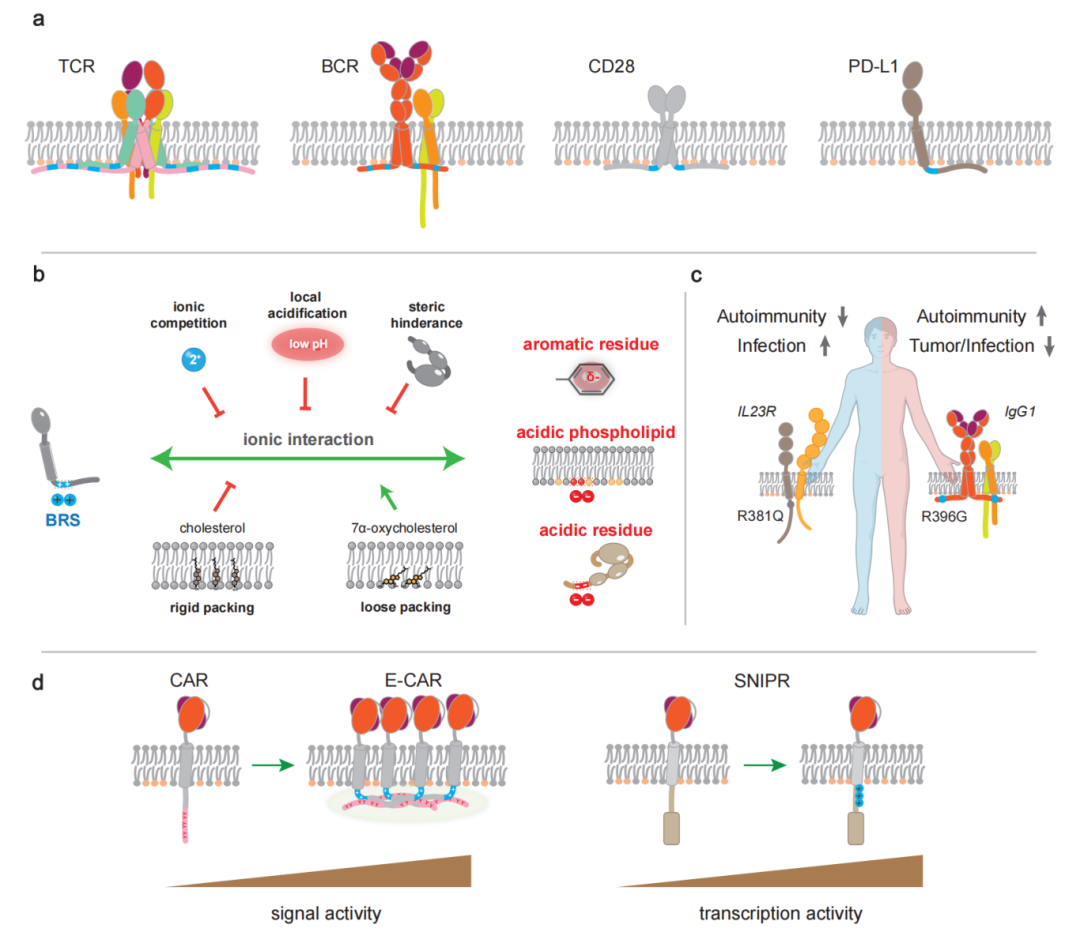
图1. 碱性残基富集序列(BRS)介导的免疫受体信号转导机制与转化应用。a,BRS(蓝色)广泛存在于各类免疫受体的胞内近膜区;b,BRS通过时空动态的近膜静电网络调控免疫受体的信号与功能;c,BRS突变与人类疾病紧密相关;d,利用BRS设计合成受体极具转化应用前景。
碱性残基富集序列(BRS)的定义与普适性
BRS通常为10个氨基酸长度,携带两个或更多的净正电荷,经常位于胞内的近膜区,也可以分布于远膜的位置。BRS通常为固有无序区(intrinsically disordered region,IDR),但在与其它分子例如酸性磷脂相互作用后,可形成二级结构。至今,抗原受体(T细胞受体、B细胞受体)、共刺激受体、共抑制受体、NK细胞受体、Fc受体、细胞因子受体等多种免疫受体的BRS已有实验报导。不仅限于免疫受体,近70%人类单跨膜蛋白的胞内近膜区均携带BRS。
BRS介导的近膜静电调控网络
实验证明,BRS可以与细胞膜和近膜区携带负电荷或π电子的脂质和蛋白质分子发生静电相互作用,而环境因子可以进一步调控这些相互作用,形成时空动态的近膜静电网络。目前已知的网络成员包括:BRS、酸性磷脂(如磷脂酰丝氨酸(PS)、磷脂酰肌醇4,5-二磷酸(PI(4,5)P2))、甾醇分子(如胆固醇、羟化胆固醇)、正电金属离子(如钙离子)、富含负电荷或p电子的膜蛋白或近膜蛋白(如LCK、p85、LAG3、PLCγ1)等。
通过近膜静电网络,BRS调节了免疫受体的磷酸化、泛素化、液液相分离、机械信号转导等过程。具体而言,BRS-脂质静电相互作用可以将免疫受体内磷酸化位点和泛素化位点屏蔽在膜内,从而限制免疫受体的基底信号和泛素化;同时,这类相互作用可以将免疫受体与信号分子PI(4,5)P2预组装,保证免疫受体处于“整装待发”状态。研究发现,BRS与PI(4,5)P2的相互作用,对免疫受体的机械响应至关重要。配体结合受体后,可能诱导受体的构象变化以及膜环境变化,引发BRS及周围序列的膜解离;随后BRS通过静电相互作用迅速招募功能蛋白质,并可能形成具有液液相分离特性的信号小体,介导免疫受体信号的触发与放大。此外,BRS膜解离后,也会诱导泛素化位点的暴露,从而产生基于泛素化的降解或者信号调控。
BRS突变与疾病的相关性
BRS相关突变导致人类疾病的例子已经被广泛报导。UniProt数据库已经包含了多个致病的BRS突变。这些突变导致碱性残基的丢失或增加,从而改变信号转导过程。例如,IL-23R R381Q与IGHG1 G396R的致病机制已经被详细研究。丢失正电的IL-23R R381Q突变会导致IL-23R信号减弱,降低免疫反应,从而降低炎症风险,但却增加感染风险。与之相反,获得正电的IGHG1 G396R突变则会导致IgG BCR信号增强,提升免疫反应,从而增加自身免疫风险,但却降低感染风险,同时在癌症病人中展示了更好的免疫治疗响应。
BRS的转化应用前景
BRS的转化研究集中在两个方面:天然免疫受体中BRS的信号调控;以及利用BRS设计合成免疫受体。例如,甾醇代谢物7α-羟基胆固醇可以削弱细胞膜脂质分子的排列致密性,帮助TCR信号亚基CD3ε的BRS更好地与膜结合,进而抑制了TCR的磷酸化。该机制已被用于TCR-T的细胞制备中,通过抑制TCR的基底信号来提升记忆细胞的比例,提高免疫治疗的长效性。另一方面,BRS已经被用于CAR-T细胞治疗中,在二代CAR分子中加入CD3ε信号区后形成的E-CAR分子具有更好的信号转导能力。其中BRS可以通过cation-π键的方式介导液液相分离的产生,帮助细胞形成更成熟高效的免疫突触,从而提升E-CAR-T细胞的抗原敏感性和长效性。值得一提的是,BRS对于另一种合成受体SNIPR(synthetic intramembrane proteolysis receptor)的高效信号转导同样至关重要。
未来方向
膜蛋白中拥有种类丰富的BRS库。但是在当前阶段,对BRS信号转导机制的理解以及应用仍然十分有限,未来还有一系列重要的问题亟需解决。例如,BRS是否可以分成多个亚类?不同的亚类是否有具有不同的信号调节模式?各类BRS突变如何导致人类疾病?如何理性操控BRS信号或者理性设计含有BRS的合成受体?这些问题的解答将极大地提升我们对免疫系统的认识并帮助免疫疗法的开发。
中国科学院深圳先进技术研究院施小山研究员为第一作者兼共同通讯作者,中国科学院分子细胞科学卓越创新中心许琛琦研究员为通讯作者。该研究获得了国家科技部,上海市科学技术委员会,中国科学院以及深圳合成生物学创新研究院的支持。
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
